In anul 1915, Twort a observat caracterul vitros (sticlos) al unor colonii de stafilococ. Dupa 2 ani (1917), dHerelle a izolat un principiu litic din filtratele de fecale de la bolnavi cu dezinterie bacilara. Cele doua descoperiri se datorau existentei unor virusuri ale bacteriilor, capabile sa lizeze celula bacteriana (bacteriofagi).
Din anul 1920, Jules Bordet si Mihail Ciuca au descris fenomenul de lizogenie (infectia simbiotica a bacteriei cu bacteriofag, fara liza celulara, insotita de modificari ale bacteriei purtatoare).
1. MORFOLOGIA SI COMPOZITIA CHIMICA A BACTE-RIOFAGULUI
O particula de bacteriofag cu morfologie tipica, cuprinde: capul si coada (ura 1).
- Capul contine un miez de acid nucleic si un invelis proteic. Miezul de acid nucleic este format din ADN cuprinzand circa 50% din greutatea uscata a bacteriofagului. Invelisul proteic (capsida) da forma poligonala a capului si este format din subunitati identice de proteina. De fapt, invelisul formeaza in spatiu o prisma hexagonala.
- Coada este partea cu care bacteriofagul se ataseaza de suprafata celulei bacteriene. Coada variaza in ceea ce priveste complexitatea structurala, de la un bacteriofag la altul (bacteriofagul T2 de Escherichia coli prezinta o coada cu complexitate foarte mare).
In general, coada are trei parti: miezul lacunar, teaca contractila si placa bazala terminala. Placa bazala terminala are forma hexagonala, purtand atasate fibrele cozii.
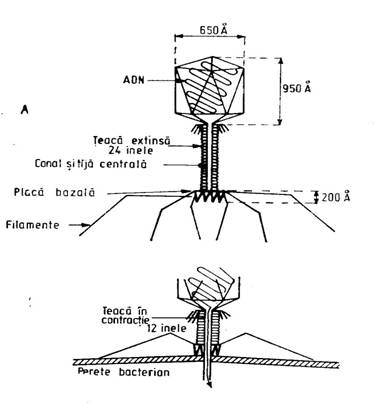
ura 1: Morfologia si stadiile functionale ale unui bacteriofag (dupa Murray, Drew, Kobayashi, 1990)
Coada bacteriofagului se poate gasi in doua stadii functionale: stadiul relaxat si stadiul contractat.
in stadiul relaxat, teaca contractila acopera aproape in intregime miezul lacunar, iar fibrele cozii nu sunt vizibile;
in stadiul contractat, teaca terminala contractila apare scurtata, miezul lacunar este vizibil pe o mai mare intindere, iar placa terminala prezinta fibrele cozii evidente.
Proteinele care formeaza capsida capului, miezul lacunar al cozii, teaca contractila si fibrele cozii sunt distincte.
In afara de forma tipica de morfologie a bacteriofagilor, pot exista si alte forme, atipice: bacteriofagi fara coada (unii bacteriofagi ADN) sau bacteriofagi filamentosi - cu morfologie variata (bacteriofagi fd in forma de bagheta, cu ADN al capului si proteinele de invelis legate in retea, insa insuficient precizata ca structura a subunitatilor componente).
1.2. FAGII ARN
Descrisi mai curand, fagii ARN (f2, MS2, R17, OB) prezinta morfologie aparte. Unui asemenea tip de bacteriofag i se descriu:
un miez de ARN monocatenar (cu greutate moleculara de 4x106 daltoni si avand o structura autocomplementara complexa, de tip tertiar);
capsida formata din circa 180 unitati (capsomere). Proteinele capsidale sunt de doua categorii si anume:
de invelis, cu greutate moleculara de 14.000 daltoni (proteina A fagica);
de maturare, cu greutate moleculara de 40.000 daltoni (ARN- polimeraza- ARN-dependenta).
Fagii ARN intervin in fenomenele de conjugare cromozomiala (fixare de fimbriile bacteriei donatoare F+), ca si in lizogenizarea unor bacterii intestinale.
2. TIPURI DE RELATIE BACTERIOFAG-BACTERIE
Bacteriofagul este un virus bacterian, a carui gazda este deci o bacterie. Atasarea bacteriofagului de suprafata celulei bacteriene depinde de existenta pe aceasta suprafata a unor receptori.
Din punct de vedere al atasarii, se disting genetic bacterii lizosensibile (care permit atasarea si intrarea bacteriofagului) si bacterii lizoresistente (care nu permit atasarea si intrarea bacteriofagului).
Lizorezistenta naturala (genetica) trebuie deosebita de rezistenta unei bacterii la infectia cu bacteriofag, datorita purtarii de catre bacterie, in intrior, a unui fag temperat (fenomen de lizogenie).
Se cunosc doua categorii de relatii bacteriofag-bacterie si anume: relatii de tip litic si relatii de tip simbiotic.
2.1. RELATII BACTERIOFAG-BACTERIE DE TIP LITIC
In acest tip de relatie, patrunderea bacteriofagului in celula bacteriana lizosensibila se soldeaza cu moartea celulei (cu unele rare exceptii), precum si cu eliberarea din celula a unor particule de bacteriofag neoformate.
Infectia fagica de tip litic implica mai multe etape si anume: adsorbtia, penetrarea, replicarea intracelulara, maturarea particulelor fagice neoformate, eliberarea particulelor fagice neoformate.
a. Adsorbtia
Datorita prezentei receptorilor pe suprafata celulei bacteriene lizosensibile, particulele de fag se adsorb pe aceasta suprafata si anume prin atasarea cozii (fibrelor cozii) pe receptori. Pe suprafata unei celule bacteriene, se pot atasa pana la 100 particule fagice.
Atasarea pe receptori se bazeaza pe proprietatea de specificitate: receptorii unei celule bacteriene sunt inalt specifici pentru anumiti bacteriofagi. Specificitatea de receptori fagici se datoreste naturii chimice a peretelui celular bacterian (dispozitia retelei de mucopeptide). De pilda, pentru bacteriofagii de Escherichia coli T3, T4, T7, receptorii se gasesc in stratul lipopolizaharidic din peretele celular, in vreme ce pentru bacteriofagii T2 si T6, in stratul de lipoproteine din perete.
Exista o competitie pe receptori intre doi fagi (celula Escherichia coli cu fagB/2 atasat, nu permite atasarea suplimentara a fagului T2).
b. Penetrarea
Odata atasat, bacteriofagul injecteaza acidul nucleic al capului in celula bacteriana, trecand din stadiul relaxat in stadiul contractat al cozii. Faptul ca acidul nucleic al capului intra in celula, iar proteina de invelis ramane in exteriorul celulei, a fost demonstrat prin marcarea acidului nucleic fagic (ADN ) cu izotopul Td3H si, respectiv, a proteinei de invelis (capsidei) cu izotopul S35.
De la aceasta regula, fac exceptie bacteriofagii filamentosi fd, care patrund in intregime in celula bacteriana (ADN + proteina).
c. Replicarea intracelulara
Replicarea bacteriofagilor ADN
Dupa cateva minute de la patrunderea acidului nucleic fagic (perioada de eclipsa), ADN-ul fagic incepe sa comande prin genele sale, in celula bacteriana gazda, sinteza unor proteine precoce (early proteins), din care fac parte enzimele necesare sintezei crescute de ADN de tip fagic. Dintre aceste enzime, cea mai importanta este o noua ADN-polimeraza (fag- dependenta), dar si altele, ca: nucleosidtrifosfatkinaza, timidil-sintetaza.
Concomitent are loc paralizarea ADN celular (oprirea sintezelor celulare codificate de ADN propriu).
In continuare, copiile de ADN fagic (rezultate din actiunea enzimelor precoce) determina, folosind aparatul de sinteza al celulei (ribozomi), formarea de neoproteine fagice tardive (late proteins), care se asambleaza in jurul moleculelor de ADN fagic, dand nastere capsidei capului si celorlalte componente proteice ale particulelor fagice neoformate.
Replicarea bacteriofagilor ARN
Cand ARN fagic intra in celula bacteriana, actioneaza el insusi ca un mesager la nivelul ribozomilor, unde se sintetizeaza mai intai o ARN-sintetaza, care asigura apoi formarea de copii de ARN fagic, care, la randul lor, codifica in ribozomi sinteza de proteine pentru formarea capsidei particulelor neoformate. O faza intermediara in replicarea bacteriofagilor ARN este formarea unui ARN dublu catenar, rezultat din lantul de ARN al particulelor fagice patrunse in celula (lant plus), legat complementar cu un lant, rezultat din actiunea ARN-sintetazei de tip fagic (lant minus).
d. Maturarea particulelor fagice neoformate
Prin asamblarea dintre moleculele de acid nucleic fagic nou formate in celulele gazda si proteinele fagice tardive, de asemenea nou sintetizate, dupa modelul codificat de acidul nucleic fagic, rezulta noile particule de bacteriofag.
e. Eliberarea particulelor fagice neoformate
La un moment dat, particulele fagice care umplu spatiul celular bacterian, produc explozia peretelui celular si membranei citoplasmatice, cu liza celulara concomitenta si cu eliberarea in mediul exterior a particulelor fagice neoformate, capabile ca, in prezenta altor celule bacteriene lizosensibile sa reia ciclul de tip litic.
Si de aceasta data, exceptia o fac bacteriofagii filamentosi fd, care trec prin membrana citoplasmatica si perete, fara a determina liza consecutiva a celulei.
2.2. RELATII BACTERIOFAG-BACTERIE DE TIP SIMBIOTIC
Fenomenul intrarii unui bacteriofag intr-o celula bacteriana, fara a determina liza acesteia si integrarea materialului genetic fagic in cromozomul bacterian, poarta numele de lizogenie.
In acest tip de relatie bacteriofag-bacterie, dupa atasarea si penetrarea bacteriofagului in celula gazda, nu se produce replicarea independenta a acidului nucleic fagic, ci acesta se integreaza in ADN celular (cromozomul bacterian), replicandu-se odata cu replicarea ADN celular (diviziunea celulei). Bacteriofagii capabili de a stabili o relatie de tip simbiotic cu bacteria purtatoare sunt denumiti profagi sau bacteriofagi temperati.
Etapele infectiei fagice de tip simbiotic sunt: adsorbtia, penetrarea acidului nucleic fagic, circularizarea ADN fagic, cuplarea ADN fagic circular cu ADN bacterian.
Bacteriile care contin in interior fagi temperati se numesc bacterii lizogene. Ele sunt rezistente (imune) la infectia cu un alt bacteriofag omolog. Acest fenomen se datoreste eliberarii in citoplasma celulei lizogene a unei substante represor, care inhiba multiplicarea unui fag litic patruns in celula.
Substanta represor actioneaza si asupra profagului existent in celula lizogena, impiedicand desprinderea acestuia din inelul comun ADN fagic-ADN bacterian si, respectiv, initierea sintezelor fagice pentru declansarea infectiei de tip litic.
Inductia reprezinta procesul prin care un fag temperat (profag) aflat intr-o celula bacteriana lizogena, trece in forma sa litica (virulenta), adica isi desprinde acidul nucleic din cromozomul bacterian si initiaza sinteze de tip fagic la nivelul aparatului ribozomul celular, cu determinarea lizei celulei si eliberarii de particule fagice virulente in mediu, capabile sa reia un ciclu de tip litic in alte celule lizosensibile. Fenomenul de inductie se datoreste inactivarii sau distrugerii substantei represor (ura 2).
Agentii capabili sa produca fenomenul de inductie (agenti inductori) pot fi variati: temperatura de 440C, iradierea cu ultraviolete, agitatia mecanica, etc. Agentii inductori ar actiona, dupa unii autori, prin determinarea in celula a sintezei unei substante inductor, care inactiveaza secundar substanta represor. Natura acestei substante inductor nu este inca cunoscuta. Pe de alta parte, apar o serie de proprietati noi ale celulei. De exemplu: tulpinile de Corynebacterium diphteriae, prin lizogenizare (purtarea unui fag temperat) devin toxigene (producatoare de toxina difterica). De asemenea, unii streptococi beta hemolitici sunt capabili de a sintetiza toxina eritrogena numai in cazul cand poarta in interior fagi temperati specifici.
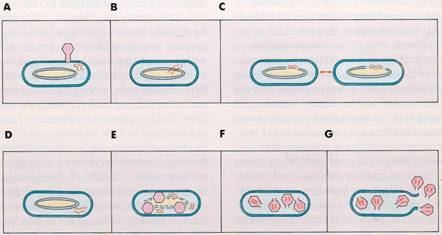
ura 2: Lizogenie-Inductie (dupa Murray, Drew, Kobayashi, 1990)
Acest fenomen de schimbare a unor proprietati ale celulei bacteriene, prin prezenta profagului, poarta numele de conversie. Intrarea fagului temperat in celula determina exprimarea unor gene celulare preexistente, represate pana atunci. Conversia lizogena difera de transductie, la care genele care caracterizeaza noile proprietati sunt cuprinse exclusiv in genomul fagului infectant.
3. IMPORTANTA FENOMENULUI DE BACTERIOFAGIE IN MEDICINA
Genetica bacteriofagilor. Bacteriofagii prezinta doua insusiri genetice importante si anume: stabilitatea tipului si rata joasa de variatie genetica. Fenomenele genetice legate de bacteriofag sunt: mutatia fagica, transductia, recombinarea fagica, restrictia si modificarea.
Mutatia fagica. Toate proprietatile unui bacteriofag sunt controlate de genele din acidul nucleic fagic. Mecanismul de mutatie genetica se aplica si la bacteriofag, ca si la bacterii, celulele animale sau virusuri animale. Deci, cu prilejul replicarii fagului in celula bacteriana, pot aparea indivizi prezentand o greseala in molecula de ADN, care confera particulei neoformate proprietati noi.
Transductia. Este fenomenul introducerii in genomul (cromozomul) bacterian a unor gene cu ajutorul materialului genetic fagic, cu aparitia consecutiva de proprietati noi ale celulei bacteriene, codificate de genele fagice introduse in ADN celular.
Recombinarea fagica. Cand pe suprafata aceleiasi celule bacteriene, se adsorb simultan doi sau mai multi bacteriofagi ADN inruditi, dar usor diferiti genetic, ambii pot infecta celula si se pot reproduce. In acest caz, unii din indivizii fagici neoformati sunt recombinati, adica poseda proprietati ale ambelor categorii de particule fagice infectante. De exemplu: daca o celula de Escherichia coli este supusa contactului cu doi fagi T2, unul care produce plaje cu dimensiuni mici, precoce, iar altul care produce plaje cu dimensiuni mari, tardiv, se obtin particule recombinate, care dau plaje cu dimensiuni mari, precoce.
Restrictia si modificarea. S-a observat ca o anumita categorie de bacteriofagi se multiplica la o rata inalta in anumite tulpini de Escherichia coli si la o rata joasa in alte tulpini apartinand aceleiasi specii bacteriene. Aceasta se explica prin faptul ca, in celula bacteriana ar exista doua enzime complementare, una modificatoare (care actioneaza asupra ADN fagic, facandu-l apt de a functiona ca matrice), alta restrictiva (care degradeaza orice forma de ADN din celula care nu a fost modificat de enzima modificatoare). In cazul infectiei cu bacteriofag a celulei bacteriene, o parte din ADN patruns din exterior nu este modificat, deci este supus degradarii, iar o alta parte, modificat fiind, nu mai este degradat si initiaza sintezele de tip fagic proprii ciclului litic. Fenomenul de restrictie se intalneste mai ales la tulpini bacteriene unde activitatea celor doua enzime este astfel coordonata, incat un numar relativ mic de molecule de ADN fagic functioneaza ca matrice pentru determinarea aparitiei particulelor fagice neoformate.
BACTERIOCINELE
Reprezinta substante de natura proteica, elaborate de anumite specii bacteriene, care au efect bactericid (litic) asupra unor tulpini de bacterii de aceeasi specie sau din specii inrudite cu a tulpinii secretoare. Actiunea bactericida depinde de existenta pe suprafata bacteriei a unor receptori specifici. Din acest punct de vedere, se disting: bacterii bacteriocino-sensibile (cu receptori) si bacterii bacteriocino-rezistente (fara receptori). Efectul bacteriocinelor este letal. De asemenea, trebuie retinut ca elaborarea de bacteriocine de catre un individ bacterian echivaleaza cu liza celulei bacteriene secretoare insasi.
Bacteriocinele prezinta o greutate moleculara relativ mare (50.000-80.000 daltoni) si un spectru antibacterian mult mai larg decat al antibioticelor. Actiunea litica a bacteriocinelor este puternica, fiind suficienta o singura molecula pentru a produce liza unei celule. Ca si in cazul bacteriofagilor, celulele bacteriene in functie de specie si tulpina, au o susceptibilitate diferita fata de bacteriocine, datorita prezentei de receptori pe suprafata peretelui celular bacterian.
Mecanismele intime ale activitatii intracelulare a bacteriocinelor comporta urmatoarele secvente: depolimerizarea ADN nuclear, inhibitia sintezei de proteine bacteriene (prin impiedicarea citirii de catre ribozom a mesajului inscris pe ARN mesager); cresterea permeabilitatii membranei citoplasmatice (cu compromiterea barierei osmotice si selective a celulei).
Din punct de vedere practic, spectrul de activitate al unei bacteriocine, elaborata de anumite specii sau tulpini bacteriene, poate fi utilizat ca indicator, el fiind caracteristic.
Metoda de testare a spectrului bacteriocinelor se numeste bacteriocinotipie (caracterizarea unei tulpini bacteriene circulante in populatia umana, pe baza spectrului litic al bacteriocinelor elaborate). Bacteriocinotipia are, deci, importanta epidemiologica, alaturi de lizotipie, pentru caracterizarea unor tulpini de Escherichia coli, Pseudomonas aeruginosa (implicate in etiologia unor infectii interioare).
Definit ca factor P sau al permeabilitatii, acesta s-a dovedit a fi reprezentat de grupul: cvercetolul, rutozida, luteolina, diosmina, hesperetolul, [...] |
Privind cu atentie ochii unui mare fumator, care obisnuieste sa locuiasca si sa lucreze in camere cu mult fum de tutun, fara aerisire, se p [...] |
Atat prin experiente pe om si pe animale, cat si din reactiile de raspuns ale bolnavilor, s-a dovedit ca VA are o actiune puternica si comp [...] |
Copyright © 2010 - 2025
: eSanatos.com - Reproducerea, chiar si partiala, a materialelor de pe acest site este interzisa!
Informatiile medicale au scop informativ si educational. Ele nu pot inlocui consultul medicului si nici diagnosticul stabilit in urma investigatiilor si analizelor medicale la un medic specialist.
Termeni si conditii - Confidentialitatea datelor - Contact